




 Current Views: 120224
Current Views: 120224
 Current Downloads: 73463
Current Downloads: 73463
Environment and Resource
ISSN Print:2707-2398
ISSN Online:2707-2401
Contact Editorial Office



Subscribe to the latest published information from SCISCAN
生物炭胶体凝聚的离子特异性效应
Ion Specific Effects on Biochar Colloid Aggregation


-
Information:
重庆科技大学化学化工学院,重庆
-
Keywords:
Biochar colloids; Dynamic light scattering; Aggregation kinetics; Critical coagulation concentration; Ionspecific effects生物炭胶体; 动态光散射; 凝聚动力学; 临界聚沉浓度; 离子特异性效应
- Abstract: Biochar, as an environmentally friendly material, is widely used in soil remediation and other fields. The aggregation and dispersion behavior of biochar colloids formed during environmental aging has an important impact on the application efficacy and potential risks of biochar in soil. The ion-specific effect refers to the different properties exhibited by different ions, especially ions of the same valence, in different systems, such as colloid aggregation/ dispersion behavior, interfacial adsorption characteristics, and particle surface potential. However, current research on the ion-specific effects of alkali metal ions on the aggregation behavior of biochar colloids is limited, and their microscopic mechanisms remain unclear. In this study, peanut shell biochar was used as the research object. The hydrodynamic diameter, critical coagulation concentration (CCC), and zeta potential of biochar colloids were measured in KNO3,NaNO3 and LiNO3 solutions at different concentrations to investigate the influence of alkali metal ions on the aggregation kinetics of biochar colloids, and thereby elucidate the alkali metal ion-specific effects and the mechanisms of their differences. The results showed that the hydrodynamic diameter of biochar colloids increased significantly with increasing electrolyte concentration. The aggregation behavior of biochar colloids in the presence of different cations displayed obvious ion specificity. Specifically, the order of the critical coagulation concentration (CCC) for biochar colloid aggregation was: Li+ > Na+ > K+. By measuring the zeta potential of biochar colloid particles in different systems, it was demonstrated that ion-specific effects dominate the differences in ionic interfacial adsorption strength during the aggregation process of biochar colloids, which is the key factor responsible for the ion-specific regulation of biochar colloid aggregation by different ions. This directly confirms the regulatory role of ion-specific effects on the aggregation behavior of biochar colloids. The findings provide a theoretical basis for evaluating the environmental fate and practical application of biochar colloids. 生物炭作为一种环境友好型材料广泛应用于土壤改良等领域,其在环境中老化形成生物炭胶体的凝聚与分散行为对生物炭在土壤中的施用功效及潜在风险具有重要影响。离子特异性效应是指不同离子,特别是同价的不同离子在不同体系中产生不同的性质差异,比如胶体凝聚分散行为、界面吸附特征及颗粒表面电位等。然而,目前针对碱金属离子对生物炭胶体凝聚行为的离子特异性效应研究较少,其微观机制尚不明确。本文以花生壳生物炭为研究对象,通过测定生物炭胶体在不同浓度的KNO3、NaNO3和LiNO3溶液下的水动力学直径、临界聚沉浓度(CCC)及Zeta电位,探究碱金属离子对生物炭胶体凝聚动力学过程的影响,进而阐明碱金属离子特异性效应及差异机制。研究发现,随着电解质浓度的增加,生物炭胶体的水动力学直径显著增大;不同阳离子作用下生物炭胶体的凝聚行为表现出明显的离子特异性,具体表现为:生物炭胶体凝聚的临界聚沉浓度(CCC)的大小顺序为:Li+>Na+>K+;通过测定不同体系下生物炭胶体颗粒Zeta电位,证明离子界面吸附强度差异由离子特异性效应主导,是造成不同离子调控生物炭胶体凝聚差异的关键因素。直观印证了离子特异性效应对生物炭胶体凝聚行为的调控作用。研究结果可为评估生物炭胶体的环境归趋及实际应用提供理论依据。
- DOI: 10.35534/er.0802037
- Cite: 魏自冕,夏金玉,刘剑伟,等.生物炭胶体凝聚的离子特异性效应[J].环境与资源,2026,8(2):273-279.
1 引言
生物炭是生物质在缺氧或限氧条件下热解制备的富碳多孔材料,具有比表面积大、表面官能团丰富、高芳香化等理化特征[1]。凭借优异的吸附性能、稳定的化学性质及环境友好特性,近年来广泛地应用于土壤改良、污染修复、碳封存与生态治理等多个领域[2-4]。已有研究表明,在农田生态系统中,生物炭可发挥多重作用[5]。例如,生物炭可以通过优化土壤孔隙结构、改良土壤物理性质及自身孔隙储水等途径,有效提升农田土壤的保水蓄水能力[6];而在石灰性农田中,长期在表层施用生物炭还能改善土壤碱性环境、促进阳离子迁移,进而加速深层次生碳酸盐的积累,最终实现深层土壤无机碳的长效固定[7]。生物炭施用于土壤后,经过干燥风化、土壤研磨、冻融、膨胀和水侵蚀等物理化学作用,会破碎形成微米、纳米级细微颗粒,并在水环境中分散形成具有高活性的生物炭胶体。环境中生物炭胶体颗粒普遍存在凝聚与分散动态平衡,该平衡过程受电解质浓度、离子价态、离子水化特征、界面极化效应等多重环境因子共同调控[8]。大量研究发现,即使价态相同的不同离子,对胶体凝聚分散行为、界面吸附特征及颗粒表面电位的调控效果仍存在显著差异,该现象即为Hofmeister效应,即离子特异性效应[9]。
离子特异性效应,最早在蛋白质凝聚试验中被发现,指同价态的不同离子,在同一体系中对颗粒凝聚分散、界面吸附、表面电位等行为产生显著差异化的客观现象[8,10]。该效应的本质是不同离子在水化半径、电子云极化程度、范德华力及短程特异性作用力存在差异,进而导致其对胶体界面性质的调控能力截然不同。对比常规宏观生物炭颗粒,生物炭胶体具有比表面积更大、表面带电官能团更丰富、界面反应活性更强等特征,对土壤溶液离子环境的变化更为敏感,其分散稳定性与凝聚行为高度依赖溶液中离子的种类及浓度[11,12]。
现阶段,针对不同价阳离子诱导生物炭胶体凝聚的动力学过程、差异化调控机制已有部分研究,而针对同价碱金属离子对生物炭胶体凝聚动力学过程的研究仍较为薄弱,特别是离子特异性效应在生物炭胶体凝聚过程中的微观作用机理有待阐明。因此,本研究采用“球
磨+离心”法制备生物炭胶体,测定Li+、Na+、K+三种碱金属离子对生物炭胶体凝聚动力学过程的影响,揭示离子特异性效应对生物炭胶体界面行为的调控机制,本研究有助于明晰生物炭胶体在土壤水环境中的沉降聚集特征,对评估生物炭长期农用的环境效应具有理论价值。
2 材料与方法
2.1 生物炭的制备
生物炭的制备原料为花生壳。花生壳生物炭烧制流程:首先用纯净水将花生壳洗净,置于70℃烘箱中烘干24 h;随后用粉碎机粉碎,过80目筛。将盛放花生壳粉末的样品舟置于管式炉间,在N2气氛下以5℃/min的升温速率升温至500℃并在此温度下热解2 h,得到本体花生壳生物炭[13]。
2.2 生物炭胶体的制备
将制备的本体花生壳生物炭在-50℃条件下冷冻24 h,使用行星球磨仪在350 r/min条件下研磨2 h(研磨球直径为3、5及8 mm,球粉比为20:1),获得球磨生物
炭[14]。取15g球磨生物炭与去离子水在500 mL的烧杯中混合后,使用超声波细胞粉碎机(JY98-IIIDN,宁波新芝生物科技股份有限公司,浙江)超声60 min,稀释使其质量分数< 1%,充分搅拌10 min。根据Stokes定律,在25℃条件下以9500 rpm的速度离心20 min获得生物炭胶体悬液[13]。
2.3 生物炭的理化性质表征分析
采用X射线衍射仪分析生物炭样品的内部晶态矿物质种类与含量;生物炭的表面化学性质采用红外光谱测定分析,光谱测定范围在400~4000 cm-1。
2.4 生物炭胶体的凝聚动力学及体系Zeta电位表征
生物碳胶体在不同电解质作用下的凝聚动力学过程,用广角动/静态激光光散射仪(BI-200SM,Brook haven,美国)测试。测试温度为25℃,散射角度为90°,数据收集时间间隔为2 min,测试时间为30 min。该仪器的工作原理是通过激光照射分散在液体中的纳米颗粒或分子,检测因布朗运动导致的散射光强度波动,并利用相关函数分析。
预先配制0.1 mol/L和1 mol/L的LiNO3、NaNO3、KNO3溶液作为母液,配置不同浓度的电解质溶液用于胶体的凝聚实验。设置以下电解质浓度梯度,LiNO3:30、80、100、200、300、400、500和600 mmol/L;NaNO3 50、80、100、200、300、400、500和600 mmol/L;KNO3 20、30、50、100、200、300、400和600 mmol/L。进行凝聚实验时依次向散射瓶中加入生物炭胶体悬液、超纯水、电解质溶液,使得待测样品总体积为10 mL。
使用纳米粒度及Zeta电位分析仪(BeNano 180 Zeta,丹东百特仪器有限公司,辽宁)测量生物炭胶体表面的Zeta电位,设置Cycle为5,重复三次。其工作原理是通过施加电场驱动带电颗粒迁移,利用多普勒效应测定电泳迁移率,结合Henry方程计算Zeta电位。测定生物炭胶体凝聚的Zeta电位时三种电解质浓度均设置为10、100、200、300、400、500和600 mmol/L。
2.5 数据处理
总体平均凝聚速率(Total Average Agglomeration Rate,TAA rate)表征胶体颗粒受布朗运动或外力碰撞聚沉的快慢,反映体系动力学稳定性。胶体凝聚的总体平均凝聚速率是根据理论基础,通过动态光散色仪连续测定胶体凝聚过程中凝聚体的粒径随时间的变化得到的,其计算公式为:

(1)
式中,为=0到任意时刻=时间段内的总体平均凝聚速率,nm·min-1;是=0到任意t时刻的平均凝聚速率,nm·min-1,为电解质的浓度,nmol·L-1;为时刻的胶体颗粒的有效粒径,nm;为颗粒的初始时的有效粒径,nm[15]。
临界聚沉浓度(Critical Coagulation Concentration, CCC)指胶体开始快速聚沉所需电解质的最低浓度。通常用于评估胶体的稳定性,CCC值越低,表明胶体在电解质溶液中越容易聚沉,胶体越稳定;反之则越难聚沉,胶体越不稳定[12]。Zeta电位是指胶体颗粒运动时,其剪切面与溶液之间的电势差。它反映了胶体颗粒表面电荷与周围分散介质之间的静电相互作用,是评估胶体稳定性和表面电化学性质的关键参数。
3 结果与分析
3.1 生物炭胶体基础性质及体系稳定性
(1)X射线衍射分析( XRD )
生物炭材料的XRD结果如图1所示,生物炭中主要存在方解石(CaCO3,23.8°、26.8°、29.3°、39.2°、43.3°和47.3°)、磷酸盐(SiP2O7,23.8°、26.8°、29.3°、44.0°和55.0°)、石墨(C,26.8°、43.3°和55.0°)的晶体结构峰。
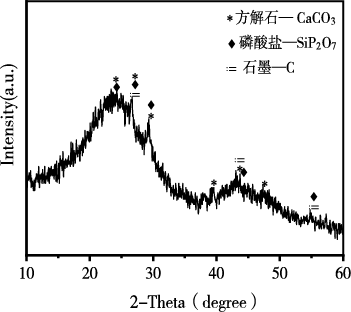
图 1 生物炭的XRD图谱
Figure 1 XRD pattern of biochar
(2)红外光谱分析(FTIR)
生物炭的红外光谱图,结果如图2所示。3416.11 cm-1处为氢键缔合的-OH伸缩振动表明生物炭表面含有丰富的含氧官能团,如羧基、羟基、羰基等,对其吸附性能和胶体稳定性有影响;2714.22cm-1和2812.04cm-1处为脂肪族C-H的伸缩振动,峰强度较弱,说明该生物炭的脂肪族结构已在热解过程中大量分解,热解程度较高,碳骨架逐渐向芳香化转变;1632.09cm-1处为芳香环C=C骨架的伸缩振动,可能叠加吸附水的H-O-H弯曲振动,这是生物炭芳香化结构的标志性峰,说明热解过程中形成了稳定的芳香碳结构;1382.55cm-1和1349.95cm-1处为O-H弯曲振动、C-O伸缩振动,进一步证明表面含氧官能团的存在。776.39cm-1和618.48cm-1为芳香环C-H的面外弯曲振动,以及硅氧键Si-O-Si的振动说明芳环已形成并带有取代基,同时生物炭中可能含有SiO2。
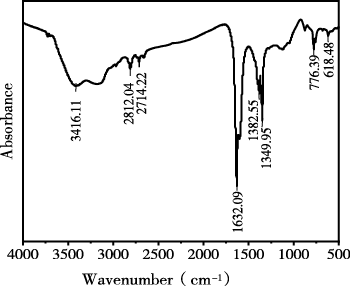
图 2 生物炭红外光谱
Figure 2 Infrared spectrum of biochar
(3)生物炭胶体粒径分布及稳定性
图3为动态光散射测得的生物碳胶体悬液的有效粒径分布以及粒径随时间的变化,结果表明生物炭胶体颗粒的有效粒径在0~1300nm的范围内,集中分布在200~400nm,平均有效粒径为251.8nm。在30 min内生物炭胶体的水动力学直径基本不变,说明我们获取的生物炭胶体是稳定的,可以进一步测定在不同种类碱金属离子及浓度下生物炭胶体凝聚状况。
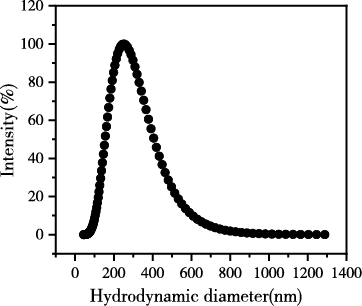
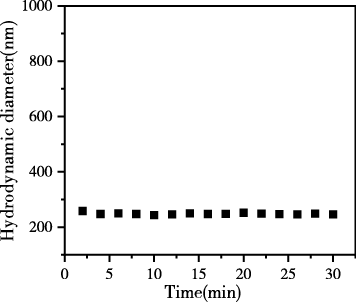
图 3 生物炭胶体颗粒的有效粒径分布和粒径随时间的变化
Figure 3 Effective particle size distribution of biochar colloidal particles and changes in particle size over time
3.2 电解质的添加对生物炭胶体凝聚动力学过程的影响
图4为添加不同浓度的LiNO3、NaNO3、KNO3电解质后,生物炭胶体的凝聚动力学过程图。结果表明,随反应时间推移,各电解质体系中生物炭胶体有效粒径均逐渐升高,说明胶体发生明显凝聚。在相同浓度相同时间下凝聚体有效粒径大小趋势表现为LiNO3 < NaNO3 < KNO3,表现出显著的离子特异性效应,粒径增长的离子特异性序列为:Li+ < Na+ < K+离子特异性。
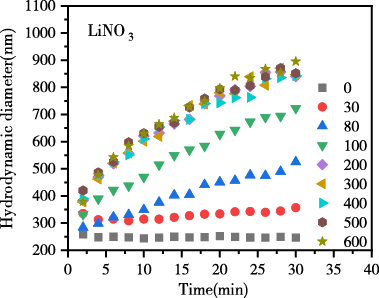
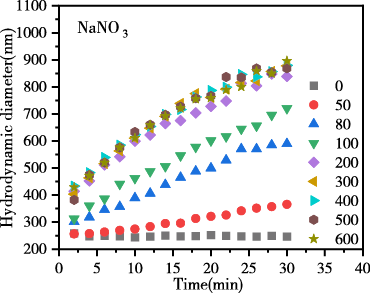
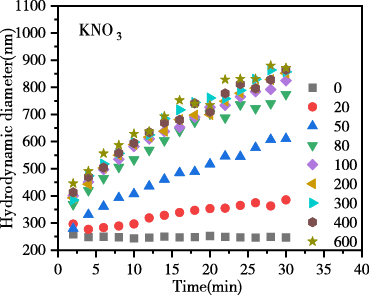
图 4 生物炭胶体凝聚过程中颗粒有效粒径随时间的变化
Figure 4 Change of effective particle size of biochar colloids over time during coagulation
3.3 离子特异性效应对总体平均凝聚速率和临界聚沉浓度的影响
基于图4中胶体粒径随时间增长的数据,可进一步得到生物炭胶体总体平均凝聚速率(TAA rates)随电解质浓度的变化规律,如图5所示。由图可知,在较低电解质浓度条件下,凝聚体粒径呈线性增长,且凝聚体在不同电解质溶液中的生长速率表现为 LiNO3 < NaNO3 < KNO3,且从图中可以看出,三种离子对应的临界聚沉浓度CCC值分别为154.46、128.14、94.41 mmol/L,离子特异性,CCC的大小为序列为:Li+ >Na+ >K+。生物炭胶体在K+、Na+、Li+离子作用下表现出明显的离子特异性效应。这种离子特异性序列反映了离子与生物炭表面含氧官能团(如羧基、酚羟基)及界面水结构的相互作用差异,进而影响胶体的聚沉动力学和稳定性。离子水合能力和电荷密度的差异对胶体稳定性的特异性影响,其中K+由于水合能力较弱,能更有效地压缩双电层并削弱颗粒表面的水化作用,导致总体平均凝聚速率最快,CCC值最低;Na+的水合能力适中,其凝聚速率和 CCC值介于 K+和 Li+之间;而 Li+因强水合作用对双电层的压缩能力较弱,且能增强颗粒表面的水化层稳定性,因此,凝聚速率最慢,CCC值最高。
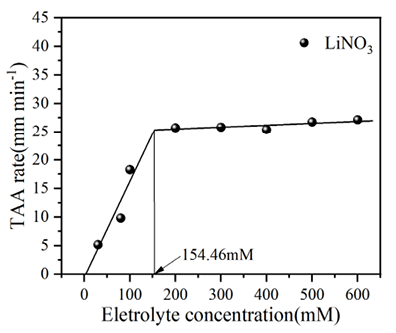
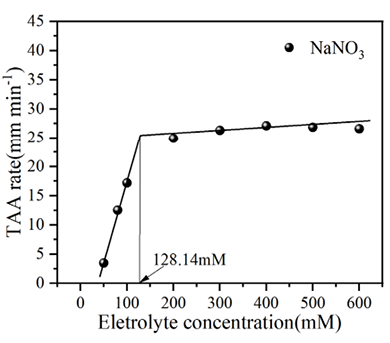
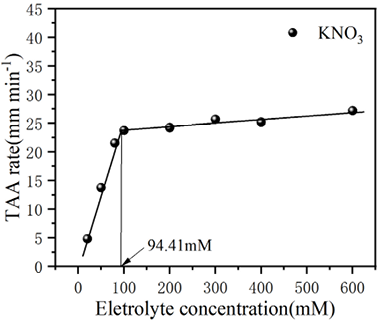
图 5 不同碱金属离子下生物炭胶体凝聚过程中的临界聚沉浓度值
Figure 5 Change of the overall average coagulation rate of biochar colloids with electrolyte concentration during the coagulation process
3.4 凝聚过程中Zeta电位随电解质浓度的变化
生物炭胶体凝聚/分散状态由颗粒间静电斥力和长程范德华引力紧密相关。Zeta电位是衡量胶体颗粒间相互排斥或吸引作用强度的重要指标,可反映胶体颗粒间静电斥力的相对大小:Zeta电位的绝对值越高,胶体间的静电斥力越大,胶体体系的稳定性越强;反之,Zeta电位绝对值越低,胶体越倾向于发生凝聚。因此,本研究不同电解质浓度对应的生物炭胶体Zeta电位,如图6所示。由图可知,所有混合胶体体系的Zeta电位均为负值,表明生物炭胶体颗粒表面均带有负电荷;随着电解质浓度的升高,Zeta电位绝对值呈现出明显下降的趋势,且不同离子作用下胶体颗粒间的Zeta电位也呈现出明确的规律: 相同电解质浓度下,Zeta电位绝对值大小遵循Li+ > Na+ > K+,即生物炭胶体颗粒Zeta电位的离子特异性序列为:离子特异性 Li+ > Na+ > K+。
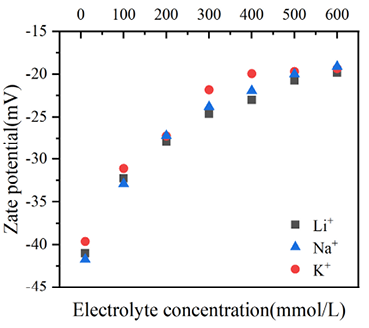
图 6 生物炭胶体凝聚过程中Zeta电位随电解质浓度的变化
Figure 6 Variation of Zeta potential with electrolyte concentration during the biochar colloid coagulation process
4 讨论
生物炭胶体凝聚与分散行为主要由胶体颗粒间微观界面相互作用力主导,其稳定与聚沉机理可通过经典DLVO理论(Derjaguin-landau-verwey-overbeek)合理解
释[16]。DLVO理论核心认为,胶体颗粒间的总作用力由范德华吸引力与双电层排斥力两部分共同决定,胶体体系的凝聚或分散状态,本质是两种作用力相互竞争、主次更替的结果[15,17]。从双电层作用机制来看,生物炭原料经高温热解后,表面富集大量羧基、酚羟基、羰基等含氧极性官能团,这类官能团在水相中易发生解离,使生物炭胶体颗粒表面整体呈现负电荷特征,颗粒表面负电荷会静电吸附水体中的阳离子反离子,在固液界面形成扩散双电层结构[18-19]。由于胶体颗粒带有同种负电荷,颗粒间会产生较强的静电排斥效应,能够有效克服范德华引力作用,阻碍颗粒相互靠近与黏附,进而使生物炭胶体长期维持分散稳定状态。而范德华引力是颗粒间固有的分子间长程作用力,作用距离相对较远,但作用强度随颗粒间距增大而快速衰减。当双电层静电斥力占据优势时,胶体保持稳定分散;一旦外界环境改变导致双电层被压缩、静电斥力显著衰减,范德华引力将占据主导,颗粒易发生碰撞、黏附并逐步形成聚集体,最终诱发胶体凝聚。
向生物炭胶体体系中引入K+、Na+、Li+碱金属离子后,溶液离子强度上升,向游离阳离子屏蔽颗粒表面负电荷、压缩双电层厚度,削弱颗粒间静电排斥力,致使胶体稳定平衡状态被打破,颗粒碰撞聚沉概率大幅提升,表现为胶体有效粒径增大、稳定性降低并发生明显凝聚。离子特异性效应进一步完善了同价离子作用机制:即便同为碱金属离子,因离子本身水化半径、水合能力、表面极化程度及界面短程作用力存在固有差异,对生物炭胶体凝聚动力学、CCC值及Zeta电位[20]都会产生截然不同的调控效果[11-12,21-22]。其中K+水合半径小、水合能力弱,更易脱去表面水化膜、贴近生物炭颗粒界面,中和表面负电荷、压缩双电层,聚沉促进作用最强;Na+水合特性居中,对胶体凝聚的诱导效果介于两者之间;Li+水化半径大、水合作用极强,外围束缚大量水分子难以靠近颗粒表面,双电层压缩效率低,对胶体凝聚的促进作用最弱,整体呈现的经典离子特异性序列规律。生物炭胶体的凝聚行为对其环境迁移性、碳封存效率及污染物吸附—运移具有重要影响,因此理解其凝聚机制对指导环境应用具有重要意义。
5 结论
对生物碳胶体凝聚速率,临界聚沉浓度及Zeta电位定量测试结果表明:随着电解质浓度升高,生物炭胶体的凝聚速率加快。不同电解质体系中,相同浓度相同作用时间下生物炭胶体的有效粒径增长速率顺序
为:Li+ < Na+ < K+,表明三种电解质体系下生物炭胶体凝聚速率呈现显著的离子特异性效应。依据生物炭胶体凝聚速率获取不同碱金属离子体系下胶体的临界聚沉浓度(CCC),且生物炭胶体凝聚的CCC值的离子特异性序列为:Li+ >Na+ > K+。而通过测定不同碱金属离子存在时生物炭胶体颗粒Zeta电位绝对值证实CCC值的离子特异性原因可能是:K+由于水合能力较弱,能更有效地压缩双电层并削弱颗粒表面的水化作用,导致总体平均凝聚速率最快,CCC值最低;Na+的水合能力适中,其凝聚速率和 CCC值介于K+和Li+之间;而Li+因强水合作用对双电层的压缩能力较弱,且能增强颗粒表面的水化层稳定性,因此凝聚速率最慢,CCC值最高离子特异性。
参考文献
[1] 徐敏,伍钧,张小洪,等.生物炭施用的固碳减排潜力及农田效应[J].生态学报,2018,38(2):393-404.
[2] 王申宛,郑晓燕,校导,等.生物炭的制备、改性及其在环境修复中应用的研究进展[J].化工进展,2020,39(S2):352-361.
[3] 陈斐杰,夏会娟,刘福德,等.生物质炭特性及其对土壤性质的影响与作用机制[J].环境工程技术学报,2022,12(1):161-172.
[4] 王重庆,王晖,江小燕,等.生物炭吸附重金属离子的研究进展[J].化工进展,2019,38(1):692-706.
[5] 高亮,李志合,王芳,等.生物炭在土壤改良中的应用进展与风险分析[J].农业工程学报,2025,41(19):19-31.
[6] Daoyuan Wang,Chongyang Li,Sanjai J,et al.Impact of biochar on water retention of two agricultural soils–A multi-scale analysis[J].Geoderma,2019,340:185-191.
[7] Yang Wang,Stephen Joseph,Xiang Wang,et al.Inducing Inorganic Carbon Accrual in Subsoil through Biochar Application on Calcareous Topsoil[J].Environmental Science & Technology,2023,57:1837-1847.
[8] Tamas Oncsik,Gregor Trefalt,Michal Borkovec,et al.Specific Ion Effects on Particle Aggregation Induced by Monovalent Salts within the 离子特异性 Series[J].Langmuir,2015,31:3799-3807.
[9] 宋冰清.生物炭微纳米颗粒的理化性质及其在环境中胶体稳定性研究[D].上海交通大学,2019.
[10] 李少博,徐英德,高晓丹,等.离子界面行为在土壤有机无机复合体形成中的作用[J].中国生态农业学报,2018,26(11):1682-1691.
[11] 杨琼芳,闫雨阳,杨东明,等.离子特异性效应对褐土和黑土胶体凝聚动力学特征的影响[J].农业环境科学学报,2025.
[12] 田锐.土壤胶体凝聚中的离子特异性效应[D].西南大学,2014.
[13] 李琪瑞,许晨阳,耿增超,等.纳米生物炭的制备方法比较及其特性研究[J].中国环境科学,2020,40(7):3124-3134.
[14] 唐尧,闫雨阳,杨琼芳,等.钾改性苹果枝条纳米生物炭的凝聚与迁移特性[J].环境科学与技术,2025,48(8):115-125.
[15] 唐嘉.“生物炭—蒙脱石”混合胶体凝聚的离子特异性效应[D].西南大学,2020.
[16] 唐嘉,朱曦,刘秀婷,等.2∶1和1∶1型黏土矿物胶体凝聚中离子特异性效应的比较研究[J].土壤学报,2020,57(2):381-391.
[17] 侯涛,徐仁扣.胶体颗粒表面双电层之间的相互作用研究进展[J].土壤,2008,(3):377-381.
[18] 丁武泉,朱启红,王磊,等.单一电解质体系下恒电荷土壤胶体扩散双电层中滑动层厚度的计算[J].土壤学报,2015,52(4):859-868.
[19] 邵珍珍,林青,徐绍辉.不同离子强度下SiO2胶体对磺胺嘧啶土壤吸附迁移行为的影响[J].土壤学报,2018,55(2):411-421.
[20] 姜军,徐仁扣.离子强度对三种可变电荷土壤表面电荷和Zeta电位的影响[J].土壤,2015,47(2):422-426.
[21] 宋孝帅,丁武泉,刘新敏,等.离子特异性效应对紫色土孔隙状况的影响机制研究[J].生态环境学报,2023,32(2):292-298.
[22] 高晓丹,寇琪,李嵩,等.黑土矿物-胡敏酸胶体复合凝聚机制:基于阳离子特异性与界面作用力的解析[J].中国环境科学,2026.



