




 Current Views: 2523776
Current Views: 2523776
 Current Downloads: 1571964
Current Downloads: 1571964
Psychology of China
ISSN Print:2664-1798
ISSN Online:2664-1801
Contact Editorial Office



Subscribe to the latest published information from SCISCAN
基于眼动追踪技术的青少年测谎实证研究
An Empirical Study on Adolescent Lie Detection Based on Eye-Tracking Technology


-
Information:
江苏警官学院,南京
-
Keywords:
Eye-tracking; Adolescents; Deception detection; Motivational intensity; Gender differences眼动追踪; 青少年; 测谎; 动机强度; 性别差异
- Abstract: Eye-tracking technology interprets visual attention and cognitive processing by recording eye movements. This technology has undergone three stages: the budding stage, the developing stage, and the mature stage, with methods continuously improving. In recent years, eye-tracking has been applied to the field of deception detection, emerging as a new technology following traditional polygraph tests. Compared with traditional methods, eye-tracking lie detection offers advantages such as simple operation, intuitive data, and non-invasiveness. Indicators including pupil diameter, fixation duration, blink frequency, and eye movement trajectory have been proven to possess strong lie detection efficacy. Given the current trend of younger age in criminal offenses, it is of great significance to study the eye movement characteristics of adolescents during deception for interrogating juvenile offenders. However, existing research has several limitations: first, a lack of eye-tracking lie detection studies specifically targeting adolescent populations; second, failure to systematically examine the influence of gender on detection outcomes; third, only comparing honest and lying groups without exploring the eye movement changes induced by different levels of motivational intensity under lying conditions. To address these limitations, this study recruited 60 adolescents (30 males, 30 females) as participants. A simulated number-writing task paradigm was adopted, with two conditions: “no motivational enhancement” and “motivational enhancement.” An Eyelink eye tracker was used to collect indicators including pupil diameter, fixation duration, blink frequency, and eye movement trajectory, aiming to reveal the eye movement characteristics of adolescents during deception and the moderating effects of gender and motivational intensity. The main findings are as follows: (1) Under the no-motivation lying condition, gender is not a key factor affecting adolescents’ eye movement patterns during deception; (2) Increased motivational intensity significantly changes the core eye movement indicators of adolescents during deception, and eye movement characteristics effectively reflect changes in cognitive load during lying; (3) The interaction between motivational intensity and gender has no significant effect on adolescents’ eye movement indicators during deception, and gender does not moderate the effect of motivational intensity on eye movement performance; (4) The experimental baseline demonstrates good homogeneity and comparability, ensuring the reliability and validity of the motivational effect analysis. 眼动追踪技术通过记录眼球运动解读视觉注意与认知加工过程。该技术经历了萌芽期、发展期和成熟期,近年来被应用于测谎领域,具有操作简单、数据直观、无侵入性等优势。瞳孔直径、注视时长、眨眼频率、眼动轨迹等指标已被证实具有较强的测谎效能。近年来,青少年涉及法律案件的比例引发关注。研究青少年说谎时的眼动特征,有助于理解其在司法问询中的行为表现。然而,现有研究存在着一些局限:一是缺乏专门针对青少年群体的眼动测谎研究;二是未系统考察性别因素对测谎结果的影响;三是未探究不同动机强度带来的眼动变化。针对上述局限,本研究以60名青少年(男女各半)为被试,采用模拟书写数字任务范式,设置“未增加动机强度”与“增加动机强度”两种条件,使用Eyelink眼动仪采集瞳孔直径、注视时长、眨眼频率及眼动轨迹等指标,旨在揭示青少年说谎时的眼动特征及性别、动机强度的调节作用。通过实验,本研究得到以下结论:(1)无动机说谎条件下,性别并非影响青少年说谎眼动模式的关键因素;(2)撒谎动机强度的提升会显著改变青少年说谎时的核心眼动指标,眼动特征可有效反映说谎过程中的认知负荷变化;(3)撒谎动机与性别的交互作用对青少年说谎眼动指标无显著影响,性别未调节动机强度对眼动表现的作用效果;(4)实验基线具有良好的同质性与可比性,保障了动机效应分析结果的可靠性与有效性。
- DOI: 10.35534/pc.0805101
- Cite: 焦语卿, 高雨菲, 冷奕帆, 范镇源, 荣欣然. (2026). 基于眼动追踪技术的青少年测谎实证研究. 中国心理学前沿, 8 (5), 678-685.
1 文献综述
1.1 眼动追踪测谎概述
眼动追踪技术,即利用特定仪器追踪并记录人眼运动轨迹的技术。一个正常人70%以上的感觉信息源自视觉,眼睛在获取信息时伴随有眼球运动,即眼动(王博,2017)。视网膜中央凹上紧密排列着分辨物体细节的视锥细胞,该区域虽仅能提供1-2度的视角,却记录了有效视觉信息的50%(Van Essen et al.,1992)。由于神经计算资源有限,人脑进化出“注意”机制,其中外显注意伴随有相应的眼球运动(Chun et al.,2011)。Just与Carpenter(1980)提出的“眼—脑一致性假说”(Eye-mind Hypothesis)证实,人的视觉和注意系统会将处于注意中心物体的光学成像聚焦于中央凹处。因此,眼动追踪技术成为解读人脑认知加工过程的重要工具。
眼动追踪测谎的发展经历了从萌芽期到成熟期的演进。19世纪末至20世纪中叶为萌芽期,研究者采用直接观察法和机械记录法初步验证了眼动行为的可记录性,但精度有限且操作侵入性强。20世纪60年代至90年代为发展期,Yarbus(1967)提出“眼动模式由任务决定”的法则,Just与Carpenter(1980)建立直接假说与眼—脑一致性假说,Kahneman(1973)的认知资源理论阐明了注意资源的有限性及其分配机制,为眼动测谎奠定了认知心理学理论基础。同期,电流记录法、电磁感应法和光学记录法等多元追踪方法相继出现,其中双浦肯野眼动仪的发明实现了高精度、无接触式眼动测量(Cornsweet & Crane,1973)。21世纪以来,眼动测谎进入成熟期,瞳孔—角膜反射(PCCR)技术成为主流,算法优化与视频记录法大幅提升了追踪精度(Duchowski,2003;Mestre et al.,2018),研究主题逐步转向眼动指标的测谎效能验证、眼动与GKT/CQT等测试范式的结合以及多模态融合(Walczyk et al.,2012;Peth et al.,2013;Gamer & Matthias,2014)。
1.2 眼动测谎的研究进展
1.2.1 眼动指标的测谎价值
近年来,眼动追踪技术因具有无损伤、生态化、操作简便等优势,逐渐进入测谎研究者的视野(任延涛 等,2016;钟少玲 等,2018)。眼动追踪通过记录被试在视觉加工过程中的瞳孔直径、注视时长、眨眼频率、眼跳及眼动轨迹等指标,可间接反映其认知负荷与情绪唤醒水平。基于“眼—脑一致性假说”(Just & Carpenter,1980),注视时间与认知加工难度直接相关;瞳孔直径对认知负荷和情绪变化高度敏感且不受主观意识控制(Heilveil,1976;Walczyk et al.,2012);眨眼频率在说谎时会出现明显的“眨眼抑制”现象(Leal & Vrij,2008;Peth et al.,2013);眼动轨迹则能综合呈现被试的扫描策略与兴趣分布(Stefan,2006;Walczyk et al.,2012)。大量模拟犯罪实验证实,犯罪者在识别与案件相关的关键信息时,瞳孔显著扩大、注视时间延长、眨眼频率降低,而无辜者则无此变化(叶小卉,2009;邬奇杰 等,2011;Lubow & Fein,1996;Webb et al.,2009)。这些结果表明,眼动指标在成人测谎中具有良好的信度与效度。
1.2.2 综合青少年特征与测谎效力的指标适用性分析
青少年期自主神经系统快速成熟,瞳孔对认知负荷的反应幅度可能大于成人(Phelps & LeDoux,2005;Beatty & Lucero-Wagoner,2000);前额叶皮层尚未完全发育,执行功能尚不成熟(Luna et al.,2004);多巴胺系统处于重塑阶段,眨眼抑制幅度可能更大且更不稳定(Jongkees et al.,2016)。这些特征既可能增强某些眼动指标的测谎敏感度,也可能引入新的干扰因素。
瞳孔直径对认知负荷和情绪唤醒高度敏感,且不易受主观意识操控,是测谎中稳定有效的指标(任延涛 等,2016)。青少年自主神经系统的快速成熟使其对认知负荷的反应更大,高动机说谎时瞳孔扩张可能更为明显。但需严格控制光线、疲劳等干扰因素。
眼动轨迹能综合反映认知负荷、情绪状态及兴趣区域分布(Stefan,2006;Walczyk et al.,2012)。青少年在场景感知中扫描策略更分散、轨迹长度更长(Luna et al.,2008;Kowler et al.,2016),说谎时轨迹凌乱性、分散度可能较成人更为显著。热力图可视化直观易懂,适合向非专业人员展示。因此,眼动轨迹适合作为青少年测谎的核心可视化指标。
说谎时出现的眨眼抑制现象在成人中敏感度达88.2%、准确率73.3%(Leal & Vrij,2008)。青少年期多巴胺系统重塑可能使眨眼抑制幅度更大,有利于检测;且眨眼频率采集方便、不易受刻意控制。但个体差异较大,适合作为辅助指标。
注视时长与认知负荷呈正相关,青少年注视稳定性较差、注视时长较成人更长(Luna et al.,2004)。但该指标可能因被试刻意回避关键信息而失真,且青少年注意力易受无关刺激干扰,导致数据波动。因此,平均注视时长适合作为探索性分析指标。
在自由观看情境下,青少年眼跳速率和幅度与成人差异不大(Bucci & Seassau,2012),但若涉及抑制眼跳的任务,青少年表现较差。本实验不要求回避特定区域,眼跳幅度可作为探索性指标。首视点易受刺激材料物理特性干扰(Charlotte et al.,2012),且实验室情境下兴趣区不明确;第二次注视时长虽记忆效应强,但对仪器精度要求较高;回视数易受其他因素影响,稳定性不足。
1.3 现有研究的不足
尽管眼动测谎已取得较为丰富的成果,但现有研究仍存在以下不足。
第一,青少年眼动测谎实证研究较匮乏。青少年生理发育的特征可能使其在说谎时的眼动变化幅度大于成人,从而有利于检测,因此需开展更多青少年眼动测谎实证研究。
第二,动机强度作为影响说谎行为的重要变量,未被系统考察。在现实测谎情境中,被测者的动机水平(如对谎言被揭穿的恐惧)存在个体差异。研究表明,高动机说谎会导致更强的认知负荷与情绪唤醒,进而放大眼动指标的变化(Walczyk et al.,2012;Proudfoot et al.,2015)。然而,现有研究,尤其在青少年群体中,动机强度如何影响眼动尚无系统研究。
第三,性别在眼动测谎中的调节作用尚不明确。叶小卉(2009)的研究未发现瞳孔反应的性别差异,但注视模式、眼动轨迹等其他眼动指标是否存在性别差异仍需进一步探索。
1.4 研究目的与假设
为弥补上述不足,研究组招募男女青少年被试,设计了适合眼动的实验范式,设置无动机与高动机两种实验条件,系统考察动机强度对青少年说谎时眼动指标的影响,并检验性别在其中的调节作用。根据已有研究,本研究最终选取瞳孔直径、眼动轨迹(含热力图)指标作为核心指标,将眨眼频率作为辅助指标,对平均注视时长、眼跳等作探索性分析。排除首视点、回视数等争议或干扰大的指标,并基于仪器局限排除第二次注视时长等指标。
研究假设如下:
(1)高动机强度会显著增大青少年说谎时的瞳孔直径、减少眨眼次数、延长平均注视时长使眼动轨迹更加凌乱分散,但对平均眼跳幅度无显著影响。
(2)性别本身对青少年说谎时的瞳孔直径、眼动轨迹、眨眼频率、平均注视时长及平均眼跳幅度均无显著影响。
(3)动机强度与性别之间无显著交互作用,即动机效应在男女青少年中表现一致。
(4)瞳孔直径与眼动轨迹(热力图)作为核心指标,能够有效区分无动机与高动机条件下的说谎反应,且热力图可直观呈现动机增强后注视热点的集中或分散变化,具有良好的可视化判别效力。
2 实验设计
2.1 被试与分组
本实验共招募被试60人,所有被试的视力或矫正视力均正常。其中男生30人,女生30人。
表 1 实验被试者构成
Table 1 Composition of experimental participants
| 被试人数/人 | 百分比 | |
| 男性被试者 | 30 | 50% |
| 女性被试者 | 30 | 50% |
| 共计 | 60 | 100% |
2.2 材料与仪器
实验图片为含数字1~7其中6个数字随机分布的图片共6张,含数字9的图片1张,均经统一处理,大小、亮度一致,全屏显示。
实验采用 Eyelink Portable Duo 型眼动仪(加拿大 SR Research 公司),刺激程序采用 Experiment Builder 软件编写,数据经 Data Viewer 导出后使用 Python 进行分析。分析方法为方差分析法,变量为“未增加动机强度”和“增加动机强度”。
2.3 实验内容
被试在纸上写10遍1~7其中一个数字以加强记忆。随后屏幕呈现6张含数字1~7随机分布的图片和1张含数字9的图片。被试需按“T”和“F”键分别表示看到和未看到自己书写的数字,同时做出说谎行为。看到数字9时按“F”。实验人员通过注视点、热力图和移动轨迹等猜测被试写的数字。
实验分两次进行。第一次为“未增加动机强度”组,第二次为“增加动机强度”组,差别在于第二组需提示被试尽量不要让实验人员猜出所写数字。
2.4 实验流程
(1)带领被试到实验室指定座位,调整位置。
(2)被试书写数字后,对眼动仪进行校准与验证。被试跟随屏幕焦点移动,不能预判注视点位置。校准合格后开始无声实验。被试观看提示语后看到三组图片,每组七张且一定含数字“9”。被试通过按“T”或“F”键判断是否有自己书写的数字,同时撒谎。每张照片停留3000ms,每组结束后休息10000ms。
(3)测试结束后,实验人员与被试一起观看数据并猜测数字,第一轮结束。
(4)第二轮“增加动机强度”实验在第一轮基础上提示被试不要被猜出数字,否则将记录为任务未完成。同样进行校准验证,提供三组照片,参数同第一轮。结束后看数据猜数字,并进行两组比较,采访被试反测谎策略。
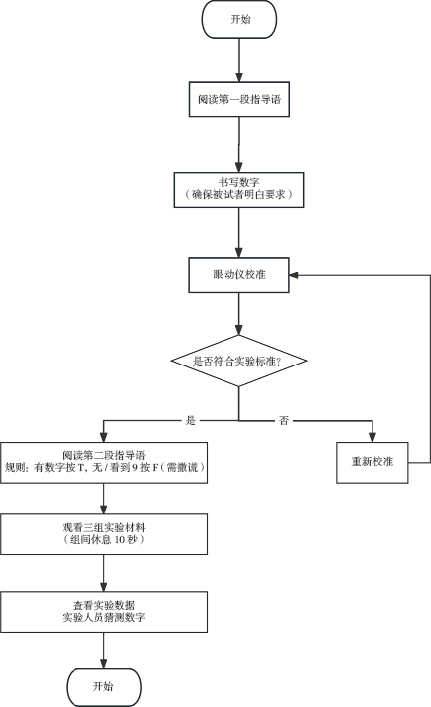
图 1 实验流程
Figure 1 Experimental procedure
3 数据分析
本实验所有眼动数据的处理与分析均采用Python语言完成,主要使用pandas库进行数据整理与清洗,scipy.stats模块实现独立样本t检验、配对t检验及卡方检验,statsmodels库中的AnovaRM函数进行重复测量方差分析。实验刺激程序采用Experiment Builder软件编写,所获得的数据经过Data Viewer软件编辑导出后,导入Python环境进行分析。
3.1 实验完成情况
本实验共招募被试60人,男女各30人。所有被试均完成无动机条件实验,数据完整有效。有动机条件下29人参与,其中24人成功匹配基线数据纳入动机效应分析,5人因无法建立个体内对照被排除。
为检验纳入样本的代表性,将24名有动机被试的无动机基线数据与仅参与无动机实验的34名被试进行基线同质性检验。检验结果表明两组在所有眼动指标上均无显著差异,确保了分析样本的同质性和可比性。在原有三项定量指标基础上,本研究进一步采集了眼动轨迹与热力图数据。
3.2 实验结果
根据眼动仪采集并导出的数据,重点对平均注视时长、平均眼跳幅度和眨眼次数三项核心眼动指标进行了统计分析,以考察不同动机条件和性别因素对眼动模式的影响。具体结果分述如下。
3.2.1 无动机条件下性别差异分析
为探究性别本身对撒谎基线眼动模式是否存在影响,对无动机条件下(n=60)男女被试的三项眼动指标进行了独立样本t检验。描述统计结果如表2所示,检验结果如表3所示。
表 2 无动机条件下男女被试眼动指标描述统计(M±SD)
Table 2 Descriptive statistics of eye movement measures for male and female participants under the no-motivation condition (M±SD)
| 指标 | 男性(n=30) | 女性(n=30) | 总体(n=60) |
| 平均注视时长(ms) | 452.60±254.98 | 349.29±203.08 | 400.95±229.03 |
| 平均眼跳幅度(°) | 5.99±1.80 | 6.32±1.55 | 6.15±1.68 |
| 眨眼次数(次/试次) | 0.69±0.59 | 0.91±0.60 | 0.80±0.60 |
表 3 无动机条件下眼动指标性别差异t检验结果
Table 3 t-test results of gender differences in eye movement measures under the no-motivation condition
| 指标 | t值 | 自由度 | p值 | Cohen’s d | 95% 置信区间 |
| 平均注视时长 | 1.67 | 58 | 0.100 | 0.44 | [-19.93,223.11] |
| 平均眼跳幅度 | -0.67 | 58 | 0.503 | -0.18 | [-1.19,0.59] |
| 眨眼次数 | -1.25 | 58 | 0.217 | -0.33 | [-0.51,0.12] |
由表3可知,在无动机撒谎条件下,男性与女性被试在三项指标上的差异均未达到统计学显著水平(所有p值均大于0.05)。这表明当未施加额外动机时,性别本身对被试撒谎过程中的眼动表现没有明显影响,男女被试眼动基线模式具有同质性。
3.2.2 动机效应分析
为考察撒谎动机强度的增强对被试眼动指标的影响及其与性别的交互作用,对成功匹配的24名被试进行了2(动机:无动机 vs. 有动机)× 2(性别:男 vs. 女)的两因素混合方差分析。描述统计结果如表4所示,方差分析结果如表5所示。
表 4 有动机组(n=24)在两种条件下的眼动指标变化(M±SD)
Table 4 Changes in eye movement measures for the motivated group (n = 24) under two conditions (M±SD)
| 指标 | 无动机(基线) | 有动机 | 变化量 |
| 平均注视时长(ms) | 378.19±180.64 | 503.40±270.44 | +125.21 |
| 平均眼跳幅度(°) | 6.18±1.37 | 5.92±1.82 | -0.26 |
| 眨眼次数(次/试次) | 0.71±0.46 | 0.54±0.52 | -0.17 |
表 5 动机效应的混合方差分析结果
Table 5 Results of mixed ANOVA for the motivation effect
| 指标 | 效应 | F值 | 自由度 | p值 | η٢ |
| 平均注视时长 | 动机 | 6.48 | (1,22) | 0.018 | 0.228 |
| 性别 | 0.22 | (1,22) | 0.647 | 0.010 | |
| 动机 × 性别 | 0.31 | (1,22) | 0.584 | 0.014 | |
| 平均眼跳幅度 | 动机 | 0.58 | (1,22) | 0.453 | 0.026 |
| 性别 | 2.46 | (1,22) | 0.131 | 0.101 | |
| 动机 × 性别 | 0.17 | (1,22) | 0.685 | 0.008 | |
| 眨眼次数 | 动机 | 4.47 | (1,22) | 0.046 | 0.169 |
| 性别 | 2.90 | (1,22) | 0.102 | 0.117 | |
| 动机 × 性别 | 0.23 | (1,22) | 0.636 | 0.010 |
方差分析结果显示,在平均注视时长上,动机主效应显著(F(1,22)=6.48,p=0.018,η٢=0.228),有动机条件显著长于无动机基线。性别主效应(F(1,22)=0.22,p=0.647)及交互作用(F(1,22)=0.31,p=0.584)均不显著。在平均眼跳幅度上,动机主效应(F(1,22)=0.58,p=0.453)、性别主效应(F(1,22)=2.46,p=0.131)及交互作用(F(1,22)=0.17,p=0.685)均不显著。在眨眼次数上,动机主效应显著(F(1,22)=4.47,p=0.046,η٢=0.169),有动机条件显著少于无动机条件。性别主效应(F(1,22)=2.90,p=0.102)及交互作用(F(1,22)=0.23,p=0.636)不显著。
进一步的简单效应分析表明,注视时长动机效应在男性中呈边缘显著(t=-2.05,p=0.068),女性中不显著(t=-1.55,p=0.147)。眨眼次数动机效应在男性中接近显著(t=1.81,p=0.100),女性中不显著(t=1.21,p=0.248)。
综合以上结果,动机强度增加能够显著延长注视时长并减少眨眼次数,反映高动机撒谎情境下认知负荷明显上升。眼跳幅度未受显著影响。性别未对动机效应产生调节作用,但简单效应分析提示动机对注视时长和眨眼次数的影响在男性中趋势更为明显。
3.2.3 基线同质性检验
为确认纳入动机效应分析的24名被试与仅参与无动机实验的34名被试在无动机条件下的眼动特征是否存在系统性差异,对两组被试的基线数据进行了独立样本t检验,结果如表6所示。
表 6 有动机组基线与仅无动机组眼动指标同质性检验
Table 6 Homogeneity test of eye movement measures between the baseline of the motivated group and the no-motivation-only group
| 指标 | 有动机组基线(n=24) | 仅无动机组(n=34) | t值 | p值 |
| 平均注视时长(ms) | 378.19±180.64 | 407.31±232.52 | -0.55 | 0.587 |
| 平均眼跳幅度(°) | 6.18±1.37 | 6.13±1.90 | -0.01 | 0.992 |
| 眨眼次数(次/试次) | 0.71±0.46 | 0.86±0.63 | -1.13 | 0.264 |
由表6可见,两组被试在三项指标上均无显著差异(所有p>0.05),说明基线眼动特征具有高度一致性,动机效应分析结果真实可靠,未受样本选择偏差干扰。
3.2.4 眼动轨迹分析
眼动轨迹图记录了被试在每张图片上注视点的移动路径和顺序。通过比较无动机与有动机条件下同一被试的眼动轨迹,发现以下典型差异。
无动机条件下,被试眼动轨迹呈现较明显的规律性和集中性,如图2所示。注视点主要分布在数字核心区域,移动路径具有较高一致性。当图片包含目标数字时,轨迹往往在该数字附近形成反复回视的闭环,或从其他区域快速跳转至该数字位置并停留较长时间。观察者可根据轨迹图较准确地推测被试所写数字。这表明无动机时被试倾向于自然地将视觉注意力集中到目标数字上,眼动模式直接反映了对关键信息的认知加工。
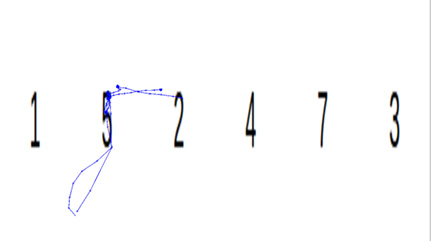
图 2 无动机的眼动轨迹
Figure 2 Scanpath under the no-motivation condition
有动机条件下,同一被试的眼动轨迹发生明显改变,如图3所示。注视点更为分散,扫描路径呈现随机或无序特征,不再围绕目标数字形成集中的回视环。许多被试故意将注视点散布到空白区域或无关数字上,试图掩盖真实关注焦点。同时眼跳幅度增大,注视点连接杂乱,难以识别稳定兴趣区。实验者无法准确判断被试所写数字。这说明当被试意识到被检测并试图隐藏真实反应时,会有意识地调整眼动策略,破坏原有规律性轨迹。

图 3 有动机的眼动轨迹
Figure 3 Scanpath under the motivation condition
3.2.5 热力图分析
热力图(基于注视时长的累积叠加图)直观反映了被试在整个实验过程中对图片不同区域的注视强度。颜色越接近红色表示注视时间越长、注视次数越多。
无动机条件下,热力图出现清晰高亮热点,精确覆盖目标数字位置如图4所示。例如当被试所写数字为“4”时,数字“4”区域呈明显红色或橙色,其他区域为绿色或蓝色。不同被试的热力图热点位置与各自目标数字一一对应,实验者可依据颜色分布直接推断目标数字。这与眼动轨迹的规律性发现相互印证,表明无动机时被试视觉注意力自然导向关键刺激。
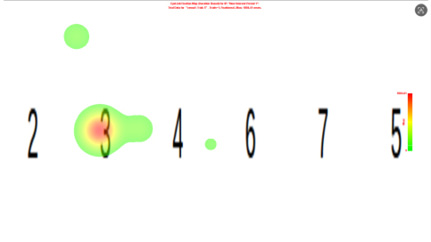
图 4 无动机的热力图
Figure 4 Heatmap under the no-motivation condition
有动机条件下,热力图原有集中热点消失,整个图片区域呈较为均匀的淡色分布,或热点随机出现在无关角落,如图5所示。尽管被试仍需识别目标数字才能正确按键,但其视觉注视被有意分散,导致目标数字区域不再获得额外注视累积。有些被试甚至反复注视干扰数字,试图制造虚假热点。因此从有动机热力图中无法分辨被试所写具体数字,热力图区分效度显著
下降。
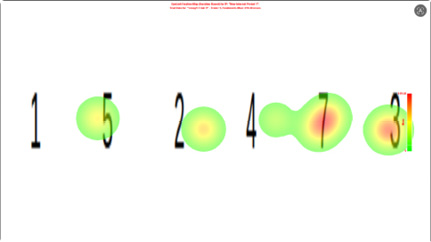
图 5 有动机的热力图
Figure 5 Heatmap under the motivation condition
如图6所示,轨迹与热力图的综合图片可更清晰对比两种条件的差异。无动机时密集轨迹线交汇于红色热点区域,形成鲜明聚焦模式;有动机时轨迹线杂乱无章,热区平散,均无法指向目标数字。这种可视化对比直观展现了动机强度对眼动模式的重塑作用。
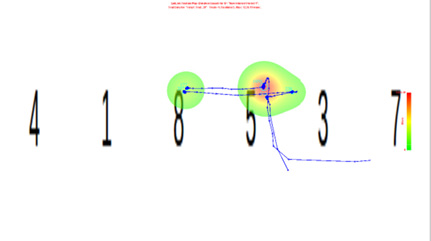
图 6 无动机眼动轨迹和热力图的复合图
Figure 6 Combined plot of scanpath and heatmap under the no-motivation condition
3.2.6 与定量指标的整合发现
眼动轨迹和热力图的定性结果与定量指标相互印证。定量分析显示,有动机条件下平均注视时长显著延长(从378.19ms增至503.40ms),但眨眼次数显著减少(从0.71次/试次降至0.54次/试次)。结合热力图和轨迹图可知,尽管被试在有动机时整体注视时间更长,但这些注视并非集中在关键目标上,而是分散到无关区域;同时,为了抑制自然注视倾向并伪装反应,被试减少了眨眼频率,这可能反映了更高的认知负荷和主动的策略控制。眼跳幅度虽有下降但不显著,而轨迹图显示有动机时眼跳路径更为混乱,这提示眼跳方向及路径的变异性可能是比眼跳幅度更敏感的测谎指标。
4 结论
在以青少年为对象的模拟说谎实验中,围绕平均注视时长、平均眼跳幅度、眨眼次数三项眼动指标,系统探究其在区分不同情境下说谎行为的有效性,同时考察性别因素与撒谎动机强度的影响。实验设置无动机、有动机两种条件,结合标准化流程采集分析眼动数据,得出如下结论。
(1)结论1:无动机说谎条件下,性别并非影响青少年说谎眼动模式的关键因素。
(2)结论2:撒谎动机强度的提升会显著改变青少年说谎时的核心眼动指标,眼动特征可有效反映说谎过程中的认知负荷变化。
(3)结论3:撒谎动机与性别的交互作用对青少年说谎眼动指标无显著影响,性别未调节动机强度对眼动表现的作用效果。
(4)结论4:实验基线具有良好的同质性与可比性,保障了动机效应分析结果的可靠性与有效性。
从实验数据来看,无动机条件下,男女被试在三项指标上均无显著差异(平均注视时长t=1.67,p=0.100;平均眼跳幅度t=-0.67,p=0.503;眨眼次数t=-1.25,p=0.217),表明性别本身并非影响无动机说谎眼动模式的关键变量。
对匹配成功的24名被试进行2×2混合方差分析,动机主效应在平均注视时长(F=6.48,p=0.018,η٢=0.228)与眨眼次数(F=4.47,p=0.046,η٢=0.169)上显著,在平均眼跳幅度(F=0.58,p=0.453)上无显著效应。有动机条件下注视时长显著延长(378.19±180.64ms vs 503.40±270.44ms),眨眼次数显著减少(0.71±0.46 vs 0.54±0.52次/试次),这与“说谎时眨眼抑制”的研究结论一致,证实高动机说谎带来的认知负荷提升会引发注视时长延长与眨眼次数减少。眼跳幅度未受动机显著影响,可能与本实验范式与刺激材料设计相关,其与说谎动机的关联仍需进一步探究。
混合方差分析显示,动机×性别交互作用在三项指标上均无显著差异(p>0.05),性别主效应也不显著,表明动机强度对眼动特征的影响规律在男女青少年中基本一致。简单效应分析提示注视时长与眨眼次数的动机效应在男性中呈接近显著趋势,在女性中不显著,提示动机效应对男性青少年影响可能更为明显,后续需扩大样本量验证。
基线同质性检验显示,24名有动机组被试与34名仅无动机组被试在三项指标上均无显著差异(p>0.05),证明基线眼动特征高度同质,样本分组不存在系统性偏差。
综合定量与定性分析,无动机条件下被试眼动轨迹具有规律性,热力图热点清晰指向目标数字;动机增强后,被试通过分散注视、扰乱扫描路径、制造虚假热点等方式隐藏真实反应,使得轨迹图和热力图无法揭露目标数字。这充分证明动机强度显著改变了青少年的眼动表征,传统眼动定性指标(轨迹、热力图)与定量指标(注视时长、眨眼次数)结合使用,可更全面评估说谎行为。
5 讨论
本研究在样本匹配和实验设计上尽量控制了潜在偏差,但仍存在若干局限性。
(1)样本匹配偏差:有动机组中仅24人成功匹配基线数据,占原始有动机组(29人)的82.8%。未匹配的5人可能导致选择性偏差。虽基线同质性检验显示匹配组与仅无动机组无显著差异,但未匹配被试可能在某些特征上存在系统性差异。
(2)性别样本不均衡:动机效应分析中男女样本量分别为13人和11人,样本偏少可能限制交互作用检测的统计功效。部分效应可能因样本量不足而未达显著水平。
(3)顺序效应:所有有动机组被试均先参与无动机实验、后参与有动机实验,无法排除顺序效应或任务熟悉度影响。未来可采用交叉设计或随机化顺序控制此类偏差。
(4)眼动指标选择:本研究聚焦注视时长、眼跳幅度和眨眼次数三项指标,未纳入瞳孔直径、注视次数、回视率等其他认知负荷敏感指标,可能限制对撒谎认知过程的多维度刻画。
(5)外部效度限制:实验在实验室环境中进行,动机操纵通过指令方式实现,与实际撒谎情境存在差距。推广至真实高动机情境时需谨慎。
综上所述,本研究在严格控制实验条件和匹配样本的基础上,揭示了动机强度对撒谎眼动行为的显著影响,具有较好的内部效度。未来可在扩大样本、优化实验设计、丰富眼动指标等方面进一步完善,以增强结论的代表性和可推广性。
参考文献
[1] 王博. (2017). 从视网膜到视皮层——视觉奥秘知多少. 科学, 69(5), 5.
[2] 任延涛, 韩玉昌. (2016). 基于眼动技术的测谎研究述评. 人类工效学, 22(2), 5.
[3] 钟少玲, 李奇光, 周建松, 张四美, 王军,郭慧娟. (2018). 眼动技术在测谎中的应用. 中国临床心理学杂志, 26(4), 3.
[4] 叶小卉. (2009). GKT测谎测试的眼动研究 (博士论文) . 浙江师范大学.
[5] 邬奇杰, 屠斌斌, 章俊龙, 宋佳. (2011). 眼动测谎指标的新探究. 科教导刊, (3), 5.
[6] Beatty J & Lucero-Wagoner B. (2012). The pupillary system. In J T Cacioppo, L G Tassinary & G G Berntson (Eds.), Handbook of psychophysiology (3rd ed., pp. 142–162). Cambridge University Press.
[7] Böhme M, Krause C, Barth E & Martinetz T. (2004). Eye movement predictions enhanced by saccade detection. In Proceedings of the 2004 Brain Inspired Cognitive Systems Conference (pp. 1-6). University of Stirling.
[8] Bucci M P & Seassau M. (2012). Saccadic eye movements in children: A developmental study. Experimental Brain Research, 222(1), 21-30.
[9] Chun M M, Golomb J D & Turk-Browne N B. (2011). A taxonomy of external and internal attention. Annual Review of Psychology, 62(1), 73-101.
[10] Cornsweet T N & Crane H D. (1973). Accurate two-dimensional eye tracker using first and fourth purkinje images. Journal of the Optical Society of America, 63(8), 921.
[11] Duchowski A T. (2003). Eye tracking methodology: theory and practice. Springer London.
[12] Gamer K M. (2013). Fixations and eye-blinks allow for detecting concealed crime related memories. International Journal of Psychophysiology.
[13] Gamer M. (2014). Mind reading using neuroimaging: is this the future of deception detection. European Psychologist, 19(3), 172-183.
[14] Heilveil I. (1976). Deception and pupil size. Journal of Clinical Psychology, 32(3), 675.
[15] Hübner W & Bennemann K H. (1973). Attention and effort. Prentice-Hall.
[16] Jongkees B J & Colzato L S. (2016). Spontaneous eye blink rate as predictor of dopamine-related cognitive function—a review. Neuroscience & Biobehavioral Reviews.
[17] Just M A & Carpenter P A. (1980). A theory of reading: from eye fixations to comprehension. Psychological Review, 87(4), 329-354.
[18] Kowler E. (2011). Eye movements: the past 25 years. Vision Research, 51(13), 1457-1483.
[19] Leal S & Vrij A. (2008). Blinking during and after lying. Journal of Nonverbal Behavior, 32(4), 187-194.
[20] Lubow R E & Fein O. (1996). Pupillary size in response to a visual guilty knowledge test: new technique for the detection of deception. Journal of Experimental Psychology Applied, 2(2), 164-177.
[21] Luna B, Garver K E, Urban T A, Lazar N A & Sweeney J A. (2004). Maturation of cognitive processes from late childhood to adulthood. Child Development, 75(5).
[22] Luna B, Velanova K & Geier C F. (2008). Development of eye-movement control. Brain and Cognition, 68(3), 293-308.
[23] Mestre C, Gautier J & Pujol J. (2018). Robust eye tracking based on multiple corneal reflections for clinical applications. Journal of Biomedical Optics, 23(3), 1.
[24] Phelps E A & Ledoux J E. (2005). Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron, 48(2), 175-187.
[25] Proudfoot J G, Jenkins J L, Burgoon J K & Nunamaker J F. (2015). Deception is in the eye of the communicator: investigating pupil diameter variations in automated deception detection interviews. IEEE.
[26] Schwedes C & Wentura D. (2012). The revealing glance: eye gaze behavior to concealed information. Memory & Cognition, 40(4), 642-651.
[27] Van Essen D, Anderson C & Felleman D. (1992). Information processing in the primate visual system: an integrated systems perspective. Science, 255(5043), 419-423.
[28] Walczyk J J, Griffith D A, Yates R, Visconte S R, Simoneaux B & Harris L L. (2012). Lie detection by inducing cognitive load. Criminal Justice & Behavior, 39(7), 887-909.
[29] Webb A K, Hacker D J, Osher D, Cook A E & Kircher J C. (2009). Eye movements and pupil size reveal deception in computer administered questionnaires. DBLP.



